
Лососи реки Варзуга
Российская Академия Наук
Кольский Научный Центр
Мурманский Морской Биологический Институт
Л.Ф.Лысенко, Е.Г.Берестовский
Лососи реки Варзуга
Мурманск
1999
Л.Ф.Лысенко, Е.Г.Берестовский. Лососи реки Варзуга
В работе рассматриваются результаты многолетних исследований по двум видам лососей, обитающих в настоящее время в реке Варзуга — семге и горбуше. Приводятся данные по численности, уровню эксплуатации, размерно-возрастному составу и возрастной структуре нерестовых стад семги реки Варзуги и основного ее притока — реки Кица. Анализируются современные представления о биологических группах атлантического лосося и их происхождении. В связи с массовым заходом горбуши в реки Варзуга и Кица, приводятся данные по численности, уровню эксплуатации и размерному составу ходового стада акклиматизанта, а также высказываются предположения по поводу возможных последствий этой экспансии.
Под редакцией к.б.н. А.Д.Чинариной
Оглавление
Динамика хода нерестовых мигрантов
Условия, влияющие на заход семги в реку
Характеристика семги р. Варзуга
Возрастная структура нерестового стада
Численность семги р. Варзуга и причины ее колебаний
О структуре чешуи семги разных биологических групп
Половая структура нерестового стада
Семга реки Кица - притока реки Варзуга
Динамика хода нерестовых мигрантов
Возрастная структура нерестового стада
Численность семги р. Кица и причины ее колебаний
Введение
В р. Варзуга, протекающей по южной части Кольского п-ова и впадающей в Белое море, в настоящее время обитает три вида лососей — семга Salmo salar L., кумжа Salmo trutta trutta L. и горбуша Oncorhynchus gorbuscha (Walbaum). Из других рыб здесь постоянно живут или заходят в эстуарную зону низовьев реки следующие виды: тихоокеанская минога Lampetra japonica Martens, обыкновенный сиг Coregonus lavaretus lavaretus (L.), европейская ряпушка Coregonus albula L., европейский хариусThymallus thymallus (L.), азиатская корюшка Osmerus mordax (Mitchill), щука Esox lucius L., плотва Rutulus rutulus (L.), елец Leuciscus leuciscus (L.), язь Leuciscus idus (L.), гольян Phoxinus phoxinus (L.), пресноводный налим Lota lota (L.), речной окунь Perca fluviatilis L., колюшки — трехиглая Gasterosteus aculeatus L. и девятииглая Pungitus pungitus pungitus (L.), ледовитоморская рогатка Triglopsis quadricornis (L) и европейская речная камбала Platichtis flesus flesus (L.).
Популяция семги р. Варзуга является крупнейшей не только на Кольском п-ове, но и среди всех других как на европейском, так и на американском континентах. На фоне других рек русского Севера, где количество атлантического лосося катастрофически снижается вследствие мощного воздействия антропогенных факторов, особенно из-за неуклонно растущего браконьерства, запасы семги р. Варзуга остаются пока достаточно стабильными, чему способствуют удаленность реки от больших населенных пунктов, ее относительная труднодоступность, отсутствие лесосплава и судоходства, как, например, на Северной Двине и Печоре.
В связи с такой уникальностью, к стаду варзугской семги необходимо бережное отношение со стороны рыбохозяйственных организаций, которые при ее эксплуатации должны руководствоваться только научно обоснованными рекомендациями. Особенно актуально это в связи с активным развитием рекреационного спортивного рыболовства и сопутствующего ему ряда негативных явлений.
Работы по акклиматизации горбуши в бассейне Белого и Баренцева морей, которые проводятся с конца 50-х годов, оказались вполне успешными. Ранее, как правило, основная часть горбуши заходила на нерест в малые реки, но в таких крупных реках, как Варзуга, фиксировался проход на нерест от нескольких десятков до нескольких тысяч рыб. Однако численность акклиматизанта в последнее время заметно увеличивается, и эта рыба начинает в массе заходить в р. Варзуга, что требует проведения тщательных исследований горбуши и выяснения особенностей ее взаимоотношений с исконным хозяином этих рек — семгой.
Кумжа, в отличие от других лососей, держится только в приустьевой эстуарной зоне, где, возможно, и нерестится на гравийных участках возле многочисленных подводных ключей. Численность ее относительно невелика, в уловах на РУЗ она встречается довольно редко, и, потому, данные по этому виду у нас практически отсутствуют. Заметим лишь, что большинство особей, попадающих на крючковые снасти, имеют массу в пределах 0,5-1,5 кг, однако в РУЗ изредка заходят крупные экземпляры массой до 3,5 кг.
Литературный обзор
Сведения по биологии атлантического лосося р. Варзуга собирались на протяжении длительного времени и, вначале, были лишь эпизодическими. Так, в работе К. М. Дерюгина (1928) со ссылкой на В. Машковцева, говорится о том, что семга, мигрирующая в р. Варзуга является наиболее мелкой, тогда как семга р. Умба — средних размеров, а самая крупная семга — в р. Нива. В работе Г. Н. Монастырского (1935), где обобщен материал по семге рек Терского берега за 1926 г., указываются размер и масса варзугской семги; возраст, в зависимости от срока пребывания рыбы в реке; абсолютный возраст дается обобщающим по всем рекам. Для анализа было взято 242 рыб, из них варзугской семги — 127 экз. Л. М. Нусенбаум (1953) изучал гематологические особенности атлантического лосося и скат молоди.
Наиболее значимые работы по изучению биологии варзугской семги были проведены М. Н. Мельниковой (1958, 1959, 1962, 1966, 1970). Опубликованные ею сведения касаются почти всех сторон биологии семги в разные периоды жизни. Характеризуя покатников, которые являются самыми мелкими для семги Терского берега, она указывала их размеры, массу и температуру воды в момент их ската в море. Работы по мечению покатников позволили установить не только пути их перемещений в пределах Белого моря, но и зафиксировать, когда на чешуе между последним речным кольцом и первым морским появляется дополнительное (Мельникова, Персов, 1968). Для взрослых особей приводятся сроки возврата в реку летней и осенней рыбы, их размеры и масса, возрастной и половой состав, плодовитость осенних производителей, процент рыб в ходовом стаде, вернувшихся в реку на повторный нерест. Показано также, что независимо от сроков входа в реку анадромных мигрантов, всегда преобладают особи в возрасте 3+1+. В то время, когда изучением биологии варзугской семги занималась М. Н. Мельникова, на реке еще не был установлен РУЗ (рыбоучетное заграждение), что, естественно, затрудняло определение численности стада и эта величина рассчитывалась на основании учета вальчаков (Мельникова, 1958). Таким образом, несмотря на трудности в сборе материала, ею была выполнена большая работа по изучению биологии варзугской семги.
На р. Варзуга работал также В. С. Михин (1958). Он обобщил и проанализировал сведения о промысле семги с конца прошлого столетия до середины пятидесятых годов нашего времени. Кроме того, он отмечал условия, которые могут влиять на количество выловленной рыбы.
Некоторые сведения по возрастному составу анадромных мигрантов семги, участию карликовых самцов в нересте, миграциях вальчаков и возврату покатников, помеченных на существовавшем ранее Варзугском рыбоводном заводе, есть в работе В. В. Азбелева (1960), который систематизировал данные по биологии семги многих рек Кольского п-ова. В дальнейшем эти работы были продолжены Э. Л. Бакштанским и М. Я. Яковенко (1974), а также В. З. Салмовым (1983).
С 1969 г. постоянные исследования на р. Варзуга стали проводить сотрудники лаборатории лососевых рыб ПИНРО, а с 1980 г. — ихтиологи Мурманрыбвода. Результаты этих исследований представлены в работах Л. Ф. Лысенко (1985, 1987, 1990а, 1990б, 1991, 1992, 1994, 1995, 1997а, 1997б), В. Г. Мартынова и Л. Ф. Лысенко (1985), М. А. Драганова, В. Г. Мартынова и Л. Ф. Лысенко (1990). Некоторые время на этой реке работал В. А. Бухтияров, который занимался исследованием и мечением “черных” и “светлых” лохов — вальчаков, разрабатывал методику идентификации варзугской семги по химическому составу чешуи (неопубликованные рукописи 1970, 1971). На основании методики О. А. Азерниковой (1964) им была разработана формула определения численности стада варзугской семги, которая, с некоторыми дополнениями, используется и в настоящее время.
Недавно издана книга «Атлантический лосось реки Варзуги», подготовленная коллективом авторов (Казаков и др., 1991). В ней, кроме собственных материалов, представлены, в частности, ранее не публиковавшиеся таблицы по характеристике анадромных мигрантов, но, к сожалению, почему-то без ссылок на их автора — Л. Ф. Лысенко. При этом содержание таблиц не всегда толкуется адекватно заключенному в них смыслу.
Материал и методика
Материал для настоящей работы был собран на фактории Усть-Кузомень на протяжении 1969-1997 гг. Рыбу на эту факторию доставляли с тоней Колониха (р. Варзуга, 12 км от устья) и Кица (р. Кица, 0,3 км от устья), где ее добывали при помощи РУЗ (рис. 1). Режим работы РУЗ и сроки его установки на протяжении этого периода менялся неоднократно. В 1969-1981 гг. РУЗ устанавливали в конце мая, и он работал в режиме «лов-пропус» 1:1. В 1982 г., с целью пропуска на нерестилища всех осенних производителей, ход которых продолжается до середины июня следующего года, лов стал осуществляться с 25 июня. С 1989 г. режим работы РУЗ стал 1:2, т.е. один день лова чередовался с двумя днями пропуска, но срок установки РУЗ оставался прежним. С 1992 г. и по настоящее время установлен пропуск на нерест 37 % производителей семги.
Поскольку есть весьма веские основания полагать, что стадо семги р. Варзуга состоит из двух практически несмешивающихся субпопуляций, одна из которых обитает в самой р. Варзуга, а другая населяет ее нижний приток — р. Кица, то мы рассматриваем их раздельно.
За весь период работы нами было исследовано 51,3 тыс. экз. варзугской семги и 12,5 тыс. экз. — кицкой, а также 315 экз. горбуши. Длина рыб измерялась по Смитту, масса — общая. У семги определяли возраст по чешуе, которую брали у рыб из 1-4 рядов над боковой линией между спинным и анальным плавниками (Major et. al., 1982; Мартынов, 1983).
Для характеристики плодовитости было отобрано 407 проб от варзугской семги и 114 проб — от кицкой. Навески икры брали в зависимости от степени зрелости икры из расчета 300-800 икринок в пробе. Подсчет икринок проводился на фиксированном материале. В связи с небольшим количеством самок тинды сбор материала по рыбам разного возраста осуществлялся в течение нескольких лет.
Для выяснения особенностей структуры речной зоны чешуи тинды и осенней семги, проведен сравнительный анализ 400 проб. Чешую измеряли под бинокуляром при увеличении х 56. Темп роста вычислен по формуле Розы Ли в модификации В. Г. Мартынова (1983).
Для характеристики хода и выявления причин колебания численности рыб использованы сведения Мурманрыбвода о вылове лососей и количестве пропущенных на нерест производителей.
Материал обработан статистически (Плохинский, 1961) и представлен в таблицах и рисунках. Таблицы, в которых содержаться сведения о длине и массе семги летней и осенней биологических групп, а также их плодовитости, являются обобщающими. Они отражают только эти параметры, и не могут быть использованы для установления соотношения полов в этих группах.
Семга реки Варзуга
Как уже отмечалось, вверх по р. Варзуга идут на нерест анадромные мигранты, которые практически не смешиваются с семгой, населяющей р. Кица.
Динамика хода нерестовых мигрантов
Ход семги в р. Варзуга начинается сразу же после освобождения ото льда, как правило, в последней декаде мая (табл. 1). Первой заходит заледка, принадлежащая к осенней биологической группе. Ее ход продолжается менее одного месяца, до середины июня. Численность заледки в уловах невелика — в среднем 1,8-2,0 % .До 1983 г. велся ежегодный учет этой рыбы, но с 1983 г. и по настоящее время заледка проходит в реку неучтенной, поскольку промысел открывается с 25 июня. Ход закройки, которая возвращается в реку после двух лет пребывания в море, продолжается с середины июня до конца июля. Этих рыб в стаде менее 1 %, а в некоторые годы она отсутствовала вовсе. С середины июня, как правило, в реку начинает идти летняя семга — тинда, ход которой завершается в первой декаде августа, но в некоторые годы этот срок сдвигается и тогда ее ход продолжается с июля до конца августа.
Таблица 1 Сроки хода разных биологических групп семги в р. Варзуга
|
биологические группы |
сроки хода |
| заледка | конец мая — середина июня |
| закройка | середина июня — конец июля |
| тинда | середина июня — август |
| осенние мигранты | середина августа — декабрь |
Количество тинды в ходовом стаде в период 1969-1995 гг. составляло 5,5-33,1 %, в среднем 18,1 %. Для нее характерны два пика подхода, приходящиеся на первую и вторую декаду июля, в среднем , соответственно, 21,8 % и 21,9 %. За период наблюдений минимальная численность тинды составила 1,4 тыс. экз., а максимальная — 18,7 тыс. экз. При этом, чем больше численность тинды в ходовом стаде, тем меньше средняя масса рыбы. Как считает Ю. П. Алтухов с соавторами (1997), увеличение доли тинды в стаде приводит, как правило, к его измельчанию, что видно из выше приведенных примеров, а также к омолаживанию. Этот вывод сделан на основании анализа динамики соотношения грилз, оцененной по данным об уловах атлантического лосося в Шотландии (Williamson, 1985). Аналогичные сведения были получены и для дальнеозерской нерки — вследствие избирательного промысла (отбирали из уловов крупную рыбу, а мелкую — отпускали), произошло увеличение доли каюрок (аналог тинды и грилз), что привело к омоложению и измельчанию стада (Алтухов, Варнавская, 1983). Интересное объяснение этому дают авторы: «Для нативных популяций нерки характерна исторически сложившаяся дифференциация менделирующих генов таким образом, что ее максимум характерен для мелких гетерозиготных самцов (каюрки, карликовые особи), минимум — для крупных самцов и средний, оптимальный уровень — для самок. В норме, благодаря селективным скрещиваниям, адаптивное преимущество имеют более гомозиготные крупные самцы … . Под воздействием промысла доля крупных более гомозиготных рыб в стаде сокращается, возрастает доля мелких, более гетерозиготных. Нарушается оптимальное соотношение полов 1:1, что ведет к уменьшению эффективной величины популяции». Поэтому, несмотря на отсутствие генетического анализа популяции варзугской семги, увеличение доли тинды в стаде должно настораживать.
После окончания хода летних мигрантов, начинается ход осенних производителей. В зависимости от условий года, наибольший подход их приходится на вторую и третью декады октября или на первую и вторую декады ноября. Так, во второй декаде октября в реку в среднем возвращается 4,9 % мигрантов (1,3 — 46,3 тыс. экз.), в третьей декаде — 6,5 % (0,7 — 45,7 тыс. экз.), в первой декаде ноября — 4,7 % (0,8 — 20,5 тыс. экз.) и во второй декаде — 3,5 % (0,6 — 8,8 тыс. экз.). К декабрю, как правило, темп подхода рыбы резко снижается. Однако в некоторые годы она продолжает интенсивно заходить в реку и в декабре, что определяется гидрологическими условиями реки.
Условия, влияющие на заход семги в реку
Количество выловленной и пропущенной на нерест рыбы в разные месяцы во многом зависит от складывающихся гидрологических и метеорологических условий — уровня и температуры воды, сроков наступления ледостава и шугования реки, направления и скорости ветра.
Сильный южный ветер во время прилива загоняет ее с пресной водой в реку и уловы возрастают, что особенно проявляется при массовом подходе рыбы в осеннюю путину, когда вслед за резким похолоданием наступает резкое потепление. И, наоборот, при сильном юго-западном ветре рыба теряет ориентир родной реки и уходит далеко на восток, за 50 км — к р. Чаваньга, где в это время уловы значительно возрастают. Такое было, например, в 1988 г., когда в конце октября ударили сильные морозы — во второй декаде было учтено 19,9 тыс. рыб, а в третьей — лишь 9,9 тыс. Только 5 ноября устье освободилось ото льда, поэтому в первой декаде ноября было учтено всего 1,3 тыс. экз. семги. Задувшие сильные юго-западные ветры отогнали семгу от устья в сторону р. Чаваньга, где промысел на морских тонях к этому времени закончился, и там снова, после большого перерыва, было выловлено около 5 тыс. рыб. В 1988 г. было учтено всего 72,5 тыс. экз. варзугской семги.
Если в октябре-ноябре, в момент подхода большого числа производителей, под действием сильных морозов появляется шуга, то рыба остается в море и только потепление в декабре способствует ее заходу. Такое наблюдалось, например, в 1969 и 1975 гг. Так, в 1969 г. в октябре было учтено 22,6 тыс. анадромных мигрантов, в ноябре — 14,4 тыс., а в декабре — 7 тыс. или 11,0 % от всей численности стада. В 1975 г. в октябре было учтено 20,5 тыс. экз., в ноябре — 40,0 тыс., в декабре — 9,9 тыс. или 10,8 % от всей численности. Промысел в 1975 и 1969 гг. продолжался до конца декабря.
Совпадение во времени негативных факторов отрицательно сказывается на численности популяции. Именно такое случилось осенью 1993 г., когда в момент пикового подхода семги (14-15 октября) наступило сильное похолодание, что вызвало большое шугование рек. В результате косяк лег под лед в районе устья р. Чаваньга и на протяжении месяца местное население собирало там погибшую рыбу, причем за один день некоторые при отливе поднимали по 150-200 экз. Как следствие, в октябре на РУЗ было учтено всего лишь 3,3 тыс. рыб. Гидрологическая и метеорологическая обстановка улучшилась только в ноябре, и последний подход семги зафиксирован в первой декаде декабря — 1,5 тыс. экз. В итоге, вместо ожидаемых 50 тыс. рыб, в реку зашло только 21,2 тыс., пропущено на нерест около 15 тыс., из них осенних мигрантов — 12 тыс. Подобная ситуация наблюдалась и ранее — в 1961 г., когда много семги погибло под шугой непосредственно в устье р. Варзуга (Лившиц, 1961).
В 1986 г после сильных осенних морозов наступало резкое потепление, да такое мощное, что вынесло лед из реки от села Варзуга (22 км от устья) далеко в море местные жители воочию смогли увидеть картину посленерестовой гибели рыбы — семгой были усеяны берега реки. Данное событие за период наших наблюдений отмечено лишь раз, да и старожилы не помнят подобного случая. Численность отнерестившегося стада 1985-1986 гг. составила около 38,5 тыс. производителей.
В 1994 г. резкое потепление в конце ноября задержало, из-за тонкого льда, установку РУЗ, поэтому в реку прошло много неучтенной рыбы, однако о хорошем ее ходе в это время можно было судить по многочисленным всплескам на середине реки в районе устья и по количеству белухи, устремившейся за этим косяком.
Возврат в реку семги на повторный нерест зависит от силы весеннего ледохода и, если этот ледоход мощный, то погибает много отнерестившейся и ослабленной рыбы, как, например, в 1997 г., в результате чего от отнерестившегося стада повторно вернулись в реку всего 0,5 % производителей.
Большое беспокойство вызывает заиливание реки и рост песчаных отмелей в низовье. Обмеление низовьев реки происходит из-за сноса песка в реку и процесс этот начался давно — тогда, когда здесь образовалось поселение Кузомень и стали вырубать лес. Но особенно интенсивно это стало происходить после 1974 г., когда в устье, за одну штормовую ночь, затянуло в песок сейнер «Челекен», который, в результате, застрял намертво и перегородил часть русла, что повлекло за собой значительное изменение гидродинамики на этом участке реки. Для сравнения приводим схематические рисунки (рис. 2), которые показывают, как выглядел этот участок реки в 1898 г. (Якобсон, 1914) и в 1978 г. В 1980 г. был поднят вопрос о закреплении берегов. Через некоторое время этим занялись сотрудники Альпийского ботанического сада КНЦ РАН, которые в с. Кузомень создали стационар, где работы проводились на протяжении 10 лет. Появились обнадеживающие результаты, но, в дальнейшем, закрепление берегов было прекращено из-за отсутствия финансирования, и возобновлено только в 1998 г., когда снова были выделены деньги на продолжение этих работ.
Обмеление реки привело к тому, что она практически перестала быть судоходной даже для маломерных лодок, а образовавшиеся обширные мелководья стали ловушкой на пути мигрирующей рыбы. Так, жители с. Кузомень не раз находили в таких ловушках по 20-30 экз. погибших рыб, причем как поздней осенью, так и весной после ледохода, ведь песок, попадающий в жабры семги на отмели, приводит к ее гибели.
Характеристика семги р. Варзуга
Основу хода варзугской семги составляют две группы рыб. Рыбы первой группы возвращаются из моря в реку летом, в ней преобладают самцы со зрелыми семенниками. Рыбы второй группы заходят в реку в осенне-зимний период, и в ней преобладают самки, которые будут нереститься только осенью следующего года после зимовки в реке.
Летняя биологическая группа. Представлена тиндой (грилз). Ее ход, как правило, длится со второй декады июня до первой декады августа. Однако, как уже отмечалось, в холодные годы эти сроки сдвигаются на одну декаду, что наблюдалось в 1994 и 1997 гг., когда первые особи тинды появились только 7 июля, а последние отмечены в конце августа. Самая поздняя поимка зафиксирована нами 4 октября — это был текучий самец. Тинда в р. Варзуга на 98,1 % представлена самцами и на 1,9 % самками (без учета возраста рыбы). Данные по длине и массы особей разного пола и возраста (табл. 2) являются обобщающими за несколько лет. Эти данные показывают, что самцы крупнее самок и пределы колебаний длины и массы у них больше.
Индивидуальная плодовитость тинды составляет 2,4-6,1 тыс. икринок (табл. 3), в среднем 4,0 тыс. икринок.
Таблица 3 Плодовитость семги р. Варзуга
|
|
|
Длина, |
Плодовитость, тыс. шт. икринок |
|||||
|
Возраст |
N, |
см |
индивидуальная |
относительная |
||||
|
|
экз. |
пределы |
средняя |
пределы |
средняя |
пределы |
средняя |
|
|
тинда |
||||||||
|
2+1+ |
44 |
44-55 |
51,3 |
2,4-5,0 |
3,7 |
1,8-3,4 |
3,1 |
|
|
3+1+ |
84 |
45-57 |
52,2 |
2,9-6,1 |
4,1 |
2,1-3,5 |
2,8 |
|
|
осенняя семга |
||||||||
|
2+1+ |
7 |
55-57 |
56,0 |
5,8-6,2 |
5,8 |
1,8-2,3 |
2,0 |
|
|
3+1+ |
153 |
58-69 |
61,0 |
6,4-9,6 |
6,9 |
1,7-3,6 |
2,6 |
|
|
2+2+ |
7 |
69-76 |
72,3 |
9,6-14,3 |
10,6 |
1,9-3,1 |
2,2 |
|
|
3+2+ |
110 |
67-86 |
67,0 |
9,0-17,9 |
11,2 |
2,6-3,8 |
3,0 |
|
|
4+2+ |
2 |
74-76 |
75,0 |
12,0-13,0 |
12,5 |
2,1-2,1 |
2,1 |
|
Осенняя биологическая группа.Вслед за ходом тинды начинается ход осенней семги (местное общеупотребительное название — «осёнка»). Сроки подхода осенних мигрантов, как и тинды, зависят от гидрологического состояния реки, Как правило, рыбы этого хода идут в реку с середины августа и до конца ноября — середины декабря, очень редко — до первых чисел января, например в 1969 г. Эти мигранты на 65-74% представлены самками.
Репродуктивный потенциал осенних производителей варзугской семги определяют в основном рыбы в возрасте 3+1+ и 3+2+ и плодовитость их составляет 6,4-17,9 тыс. икринок (табл. 3), в среднем 8,7 тыс. икринок.
Осенью в р. Варзуга входят и повторно нерестующие особи. Возврат их на повторный нерест во многом зависит от условий, складывающихся на нерестилищах, и от условий ледохода.
Наиболее ранний возврат повторно нерестующих особей отмечен в середине июня 1984 г. Это была самка длиной 51 см и массой 1,3 кг, в возрасте 3+1+Sm+, т.е. после нереста в 1983 г. и зимовки в реке, скатившись в море в мае 1984 г., она, уже в июне, вернулась в реку, приобретя нерестовую марку. У нее от первого нереста в ястыках осталось немного невыметанной икры.
Таблица 4 Длина и масса повторно нерестующих особей семги в р. Варзуга
|
Возраст |
N, |
Длина, см |
Масса, кг |
||
|
|
экз. |
Пределы |
Средняя |
Пределы |
Средняя |
|
2+1+Sm+ |
27 |
63-77 |
68,5 |
2,4-4,8 |
3,9 |
|
2+2+Sm+ |
2 |
74-82 |
78,0 |
2,5-5,6 |
5,1 |
|
3+1+Sm+ |
320 |
63-74 |
74,1 |
2,4-6,4 |
4,4 |
|
3+2+Sm+ |
19 |
73-90 |
78,7 |
4,5-9,6 |
5,1 |
|
4+1+Sm+ |
35 |
68-81 |
73,2 |
3,8-5,9 |
4,2 |
|
4+2+Sm+ |
4 |
62-74 |
76,5 |
2,4-4,2 |
4,0 |
|
3+1+Sm+1 |
4 |
61-76 |
71,0 |
2,4-4,7 |
3,9 |
|
3+2+Sm+1 |
2 |
72-83 |
77,5 |
4,5-6,4 |
4,5 |
|
4+1+Sm+1 |
1 |
72 |
72,0 |
4,5 |
4,5 |
Для повторно нерестующих особей варзугской семги характерно то, что основная их часть возвращается в реку в год ската и зимует в ней до следующего года. Однако изредка встречаются и такие рыбы, которые зимуют в море и только летом следующего года заходят в реку со зрелыми — III-IV стадии — половыми продуктами. По внешнему виду они не отличаются от остальной семги и о первом их нересте можно узнать лишь по структуре чешуи, на которой отмечен период пребывания в море в виде большого годового прироста, и этот возраст обозначается 3+1+Sm+1. Плодовитость таких рыб при длине 72 см составляет 12,5 тыс. икринок, а при длине 83 см — 19,0 тыс. икринок.
Представленные данные о повторно нерестующих особях (табл. 4) свидетельствуют о том, что на нерест чаще всего возвращаются рыбы в возрасте 2+1+Sm+, 3+1+Sm+, 4+1+Sm+, 3+2+Sm+.
Чем больше численность стада, тем выше посленерестовая смертность рыбы. Так от нерестового стада 1981-1982 гг., численностью 12,9 тыс. экз., на повторный нерест пришло 6,6%, а от нерестового стада 1987-1988 гг. численностью 85, 0 тыс. экз. — только 1,8 % (Лысенко, 1994).
О повторном нересте самок, которые зимуют до следующего нереста в реке, можно судить не только по наличию на чешуе нерестовой марки и дополнительным точкам на жаберных крышках, но и по остаткам в ястыках резорбированной икры в виде черных точек. У семги, вернувшейся в реку на повторный нерест после зимовки в море, такая икра отсутствует.
Возрастная структура нерестового стада
Основу нерестового стада составляют особи с абсолютным возрастом от 3+ до 6+лет, из них более 70 % приходится на рыб в возрасте 3+1+ (табл. 5). Особи с абсолютным возрастом 7+ встречаются крайне редко. Такая семга, в частности, была поймана в сентябре 1996 г. и имела длину 180 см, массу — 21 кг, возраст 4+3+.
Тинда, помимо основной возрастной группы 3+1+, представлена также рыбами в возрасте 2+1+ и 4+1+, доля которых варьирует по годам. Крайне редко встречается особи в возрасте 5+1+ — их длина составляет 45-53 см.
Основная масса осенних мигрантов также представлена рыбами в возрасте 3+1+. С увеличением численности стада доля их повышается, и в годы максимальной численности (1975, 1987) она достигает более 80 % в ходовом стаде. Помимо основной возрастной группы 3+1+. осенние мигранты представлены и другими возрастными группами — 2+1+, 4+1+, 2+2+ и 3+2+. Рыбы в возрасте 3+3+ и 4+2+ встречаются значительно реже, и, совсем редко, особи в возрасте 5+1, длина которых колеблется в пределах 48-59 см. При абсолютном возрасте, равном 6+, как и у рыб возрастных групп 3+3+ и 4+2+, они отличаются от них длиной и массой, потому что покатники имели разную длину. Так, смолты в возрасте 5+ скатываются из реки при длине 12,8-,15,4 см, изредка до 17,2 см, а в возрасте 3+ и 4+ — при длине 8-10 см. Чем больше длина покатника, тем ниже темп роста рыбы в море.
Известно, что у атлантического лосося встречаются особи, возвращающиеся в реку в год ската на стадии смолта, но они настолько редки в популяции, что нам, за весь период исследований, удалось обнаружить лишь 5 таких рыб. Это были самки, выловленные с августа по октябрь, которые имели длину 33 -42 см, возраст 3+0+ и 4+0+, яичники II стадии зрелости. У самки длиной 42 см были просчитаны ооциты. Абсолютная плодовитость составила 2,6 тыс. икринок. Известно, что в реку в год ската возвращаются также и самцы (Азбелев, 1966), но нам они не попадались.
Возраст рыб, которые вернулись в реку в год ската, к моменту нереста будет равняться четырем годам:3+0+1+ (зимовка в реке до нереста). Сравнение абсолютного возраста этих рыб с абсолютным возрастом летней и осенней семги на момент нереста позволило предположить, что самки, вернувшиеся в реку в год ската, вырастают из той же молоди, что и тинда. Сделать такой вывод позволили не только сравнительный анализ возраста, но и анализ размерных показателей. Так, минимальные размеры самцов и самок тинды составляют, соответственно, 37 и 42 см, а осенней семги в возрасте 3+1+ — соответственно, 48 и 50 см. Достоверно известно, что тинда — это бывшие карликовые особи (Schiffer, 1971; Митанс, 1973; Larsson, Svensson, 1974; Лейзерович, Мельникова, 1979; Myres et. al., 1986). Исходя из этого, мы склонны относить самок, вернувшихся в реку в год ската, также к бывшим в реке половозрелыми карликовым особям. Заметим, что в настоящее время появляется все больше сведений о нахождении в естественных популяциях карликовых самок (Prouzet, 1981; Bgliniere, Massie 1985; Hindar, Nordland, 1989). На Кольском п-ове, в частности, карликовая самка была поймана сотрудником ПИНРО М. Я. Яковенко в июле 1995 г в р. Ура, впадающей в Баренцево море на Западном Мурмане. Она имела длину 19,6 см, массу 86,0 г, возраст 4+, масса гонад составила 0,45 г.
Однако эти данные по возрасту являются обобщающим и не позволяют судить о роли каждого поколения в воспроизводстве. Для подробного анализа возрастной структуры в качестве примера использованы сведения о нерестовом стаде за 1994 г. (табл. 6), где указываются год появления потомства, сроки пребывания в реке, длительность нагула в море и год возврат в реку, где они зимуют и находятся в течении года до нереста. Таким же образом характеризуется молодь, созревающая в реке на стадии пестрятки — карликовые особи, которые становятся затем тиндой. Данные о рыбе, принимающие участие в нересте повторно, представлены несколько иначе. Отнерестившись первый раз, производители, скатываются в море весной следующего года, в тот же год заходят в реку, чтобы, перезимовав, снова принять участие в нересте.
Анализ возрастной структуры нерестового стада, выполненный по такой методике, показывает, что в нересте принимают участие рыбы 8-ми поколений, составляющих 12 возрастных групп рыб осенней биологической группы, и по 3 возрастные группы тинды и карликовых особей. Исходя из этого удалось установить, что основные производители нерестового стада могут быть оплодотворены своими же карликовыми особями, представленными в стаде тиндой в возрасте 4+1+. Такой особенности размножения варзугской популяции семги нельзя было заметить при прежнем способе анализа возрастной структуры нерестового стада. Кроме того, наглядно показано, что лососи, принадлежащие к более раннему поколению и к более позднему, представлены меньшим количеством возрастных групп, чем поколения, которые концентрируются возле основных производителей. При этом своими оплодотворяющими качествами они сильно отличаются друг от друга. Так, особи, родившиеся от производителей 1983 г. возврата, представлены в новом нерестовом стаде повторно нерестующими особями, а рыбы, родившиеся от производителей 1990 г. возврата — карликовыми особями. Это означает, что качество половых продуктов у них разное, что подтверждается экспериментами по изучению влияния качества производителей на качество выращиваемых на рыбоводных заводах молоди лосося (Песлак, 1967). Результаты зкспериментов показали, что физиологическая ценность спермы и икры у молодых рыб выше, чем у особей старшего возраста, половые продукты которых мало пригодны для получения полноценной молоди. Кроме того, было экспериментально доказано (Thorpe et. al. 1983, Thomaz et. al. 1997)., что при оплодотворении самок карликовыми самцами и самцами тинды рождается больше карликовых самцов, чем при оплодотворении проходными самцами осеннего хода. Возможно, что именно по этой причине так сложна нерестовая структура стада семги и так трудно прогнозировать численность тинды.
Численность семги р. Варзуга и причины ее колебаний
Численность ходового стада семги р. Варзуга сильно колеблется по годам и находится в пределах 18-140 тыс. экз. (табл. 7).
Для объяснения спадов и подъемов численности следует обратить внимание на две особенности. Во-первых, как уже отмечалось, нерестовое стадо на 65-74 % представлено самками и на 70-75% — рыбами в возрасте 3+1+, а в годы большой численности количество рыб с абсолютным возрастом 4+ может достигать более 80%. Во-вторых, в отличие от популяций атлантического лосося большинства других рек Кольского п-ова, основная масса рыб в которых возвращается в реку в весенне-летний период и нерестится в год возврата, ходовое стадо варзугской семги формируют в основном рыбы осенней биологической группы, нерестящиеся осенью следующего года. По этой причине отсчет ожидаемой численности стада семги мы проводим не с момента нереста, а с момента захода в реку осенней семги. Этот метод был применен именно к лососю р. Варзуга из-за несовпадения сроков возврата основной группы производителей и сроков их нереста. Соответственно, надо знать, сколько рыбы вернется в реку от основных производителей, потому что в тот год, когда происходит их нерест, в реку заходят рыбы уже другого поколения.
.
Таблица 7 Характеристика стада семги р. Варзуга и уровень его эксплуатации за период 1975-90 гг.
|
Год |
Числен-ность, |
Вылов, |
Средняя масса особи, |
Соотношение полов, % |
Соотношение рыб, % |
|||
|
|
тыс. экз. |
тыс. экз. |
ц |
кг |
самцы |
самки |
тинда |
осенняя |
|
1975 |
90,0 |
44,9 |
1251 |
2,8 |
37,5 |
62,5 |
8,0 |
92,0 |
|
1976 |
57,7 |
28,6 |
855 |
3,0 |
33,8 |
66,2 |
8,9 |
91,1 |
|
1977 |
35,8 |
17,7 |
469 |
2,7 |
32,5 |
67,5 |
11,6 |
88,4 |
|
1978 |
18,8 |
9,2 |
246 |
2,7 |
41,2 |
58,8 |
12,3 |
87,7 |
|
1979 |
41,4 |
20,5 |
524 |
2,6 |
40,5 |
59,5 |
10,0 |
90,0 |
|
1980 |
44,0 |
21,8 |
707 |
3,2 |
31,7 |
68,3 |
3,0 |
97,0 |
|
1981 |
29,2 |
16,1 |
490 |
3,1 |
36,1 |
63,9 |
11,1 |
88,9 |
|
1982 |
24,8 |
13,4 |
461 |
3,4 |
30,4 |
69,6 |
3,2 |
96,8 |
|
1983 |
60,0 |
29,9 |
954 |
3,2 |
33,8 |
66,3 |
4,9 |
95,1 |
|
1984 |
40,2 |
19,8 |
634 |
3,2 |
28,8 |
71,2 |
12,2 |
87,8 |
|
1985 |
47,8 |
23,5 |
701 |
2,9 |
30,0 |
70,0 |
22,7 |
77,3 |
|
1986 |
70,9 |
35,1 |
955 |
2,7 |
20,7 |
79,3 |
15,8 |
84,2 |
|
1987 |
137,0 |
50,7 |
1612 |
3,2 |
29,5 |
70,5 |
9,4 |
90,6 |
|
1988 |
72,1 |
26,4 |
801 |
3,0 |
25,3 |
74,7 |
8,2 |
91,8 |
|
1989 |
65,1 |
24,4 |
709 |
3,0 |
35,0 |
65,0 |
22,9 |
77,1 |
|
1990 |
39,5 |
14,1 |
499 |
3,5 |
30,6 |
69,4 |
19,7 |
80,3 |
Поскольку основу нерестового стада составляют особи, мигрирующие в реку в осенний период и имеющие возраст 3+1+, можно заключить, что с момента захода в реку основных производителей нерестового стада и до возвращения из моря большинства их потомков проходит 6 лет. Этот цикл оборачиваемости можно проиллюстрировать на конкретном примере. Осенью 1987 г. в реку вернулись основные производители, которые остались зимовать в реке. Осенью следующего, 1988 г., они отнерестились вместе с тиндой другого поколения, вернувшейся в реку летом этого года, и карликовыми особями. Весной 1989 г. появились мальки в возрасте 0+, в 1990 г. им было 1+, а в 1991 г. — 2+. В 1992 г. большинство из них в возрасте 3+ скатилось из реки в море. В 1993 г. основная часть рыб, пробыв в море один год, вернулась в реку в возрасте 3+1+ двумя типами хода — летним и осенним. Таким образом, от захода в реку основных производителей и до возврата от них основной массы новых производителей проходит шесть лет. Аналогичные суждения были высказаны В. В. Азбелевым (1968), который также отмечал, что среди рыб, заходящих в реки Белого моря, всегда преобладают рыбы осеннего хода в возрасте 3+1+, и при средних условиях значительная часть поколений, воспроизводящихся в реках, мигрирует на нерест через шесть лет после захода производителей.
На основании предлагаемого толкования жизненного цикла варзугской семги был проведен анализ численности стада. Динамику хода в период с 1945 по 1994 гг. мы рассматривали как последовательный переход одного поколения осенней семги в другое, которые и составляют отдельные воспроизводящие линии
1945-1951-1957-1963-1969-1975-1981-1987-1983 — 1линия
1946-1952-1958-1964-1970-1976-1982-1988-1994 — 2 линия
1947-1953-1959-1965-1971-1977-1983-1989-1995 — 3 линия
1948-1954-1960-1966-1972-1978-1984-1990-1996 — 4 линия
1949-1955-1961-1967-1973-1979-1985-1991-1997 — 5 линия
1950-1956-1962-1968-1974-1980-1986-1992-1998 — 6 линия
Очевидно, именно с наличием и состоянием этих линий в популяции и связаны колебания численности стада. Анализ динамики уловов (рис. 3) с выделением шести воспроизводящих линий позволили установить закономерности, проявляющиеся в чередовании спадов и подъемов, как в отдельных линиях, так и в стаде в целом. При этом величины спадов и подъемов в каждой из линий не равноценны друг другу, что связано не только с различной степенью эксплуатации стада, но и с гидрологическими условиями реки в осенний период (Лысенко, 1994).
В период 1945-1965 гг. — до введения правил концентрированного лова — варзугская семга интенсивно облавливалась не только в реке, но и на многочисленных морских тонях. Между тем, мечение семги на трех морских тонях (Трухинская, Пахомовская, Шерстинская) показало, что большая часть выловленных здесь рыб приходится именно на семгу варзугской популяции (Мартынов, Лысенко, 1985), и становится понятным, почему предшествующий нерациональный промысел так сильно сократил численность этого стада. С целью сохранения природных запасов семги, В. В. Азбелевым были предложены правила концентрированного лова, обеспечивающего контролируемый пропуск производителей на нерестилища, которые начали действовать с 1966 г.
Установление контролируемого пропуска рыбы способствовало нарастанию численности варзугской популяции семги, позволило вести учет пропущенной на нерест рыбы и получать достоверные данные по периодичности спадов и подъемов в каждой из шести воспроизводящих линий, что является основой для прогнозирования. В частности, оказалось, что в разумно регулируемой популяции семги, закономерны колебания численности по типу «с большой на малую и с малой на большую». Это позволяет заранее прогнозировать динамику вылова рыбы.
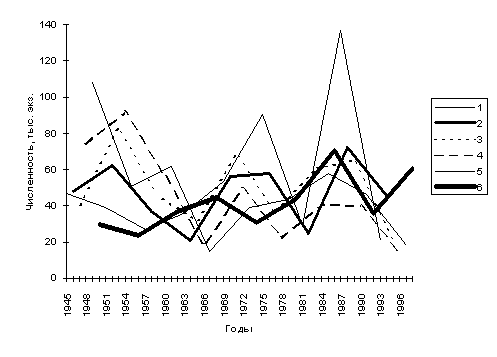
Рис. 3. Динамика численности ходового стада семги р.
Варзуга за период 1945-1998 гг.
Выяснению причин, влияющих на периодичность уловов семги, посвящено ряд исследований. И. Б. Бирман (1969), основываясь на суждениях Л. С. Берга, полагал, что чередование урожайных и неурожайных поколений семги бассейна рек Баренцева и Белого морей свойственны 8-11-летние циклы. Причину этих колебаний он связывал с солнечной активностью, а поколения, дающие минимальные и максимальные уловы, определял, отнимая 5 лет от года улова. Однако, принимая во внимание тот факт, что сроки пребывания молоди в разных реках различны, так же как и сроки пребывания рыбы в море, трудно согласиться с таким способом деления семги на урожайные и неурожайные поколения. Так, например, молодь живет в реках бассейна Баренцева моря в целом дольше, чем в реках бассейна Белого моря. Количество рыб, нагуливающихся в море два года у баренцевоморской семги значительно выше, чем у беломорской, поэтому годы с минимальными и максимальными уловами у них не могут совпадать. Поэтому нужен дифференцированный подход к анализу динамики вылова и прогнозу численности для популяций семги Баренцева и Белого морей. В частности, для определения максимальной и минимальной численности популяции семги р. Варзуга подход И. Б. Бирмана неприемлем, поскольку здесь стадо мигрантов имеет двойное происхождение: оно состоит из рыб весенне-летнего хода, нерестящихся в год захода в реку, и рыб осеннего хода, принимающих участие в нересте только на следующий год. Если придерживаться метода И. Б. Бирмана, то для прогнозирования ожидаемой численности следует брать только часть производителей, т.е. рыб весенне-летнего хода. Численность их мала по сравнению с количеством семги осеннего хода, и, соответственно, они не могут внести решающего вклада в воспроизводство стада. Кроме того, как уже указывалось, в потомстве, полученных от самцов тинды, высок процент карликовых особей.
Для прогнозирования урожайных и неурожайных поколений неприемлем также способ, используемый В. В. Покровским (1966), где отсчет численности нового поколения ведется с момента его рождения, т.е. не учитывается время нахождения производителей в реке до нереста. В результате, нарушается цепь непрерывного перехода рыб одной генерации в другую, что, естественно, искажает связи и закономерности, поскольку величина потомства не соотносится с численностью родительского стада.
Итак, предложенный нами подход к оценке структуры и состояния популяции варзугской семги, позволил объяснить причины закономерных колебаний численности стада и повысить обоснованность прогнозов.
О структуре чешуи семги разных биологических групп
Определяя возраст семги, мы заметили, что рыбы в возрасте 3+1+, возвращающиеся в реку в разное время, отличаются друг от друга структурой речной зоны чешуи. Эти различия между ними являются постоянными, повторяющимися из года в год. Причину этого явления мы стремились установить путем проведения сравнительного анализа и использования литературных данных.
При сравнении чешуи семги летнего и осеннего хода были выявлены различия в структуре речной зоны на втором (1+) и третьем (2+) годах жизни (Лысенко, 1997). Так, у большинства особей тинды обоих полов в возрасте 3+1+ приросты на третьем году жизни больше, чем на втором, а у семги осеннего хода такого же возраста годовые приросты на втором и третьем годах, как у самок, так и у самцов, практически одинаковы (табл. 8).
Таким образом, в результате проведенного исследования установлены такие различия в структуре чешуи, которые позволяют достоверно различать принадлежность рыб к той или иной биологической группировке (Л. Ф. Лысенко, а. с. 1655416 СССР, МК5А01К61/100.-14684758/13). Бюлл. 22 от 12.06.91). Эти различия постоянны, как стабильно и то, что тинда представлена практически одними самцами, а у осенней семги преобладают самки. Мы связываем это с существованием у атлантического лосося двух типов молоди — половозрелых карликовых особей и неполовозрелых пестряток. Л. С. Берг (1961) считал, что такая молодь рождается от одних родителей. Так, первые в преднерестовый период начинают усиленно питаться, в результате чего у них повышается темп роста (Лейзерович, 1973; Jones, 1949), что отражается на чешуе в виде большого прироста и большего расстояния между склеритами, чем за предыдущий год. Такие же особенности в структуре чешуи (табл. 9)
характерны и для карликовых самцов варзугской семги (Мартынов, 1983). У неполовозрелых пестряток все годовые приросты, как правило, одинаковы (Митанс, 1973; Казаков, 1979; Larsson, Svensson, 1974; Bailey et. al., 1980; Myres et. al., 1986). Данное явление свойственно и для тихоокеанских лососей (Горшков, Горшкова, 1977; Падецкий, Флоряк, 1983).
Поскольку после пребывания в море структура речной зоны чешуи не меняется, мы сопоставили структуру речной зоны чешуи тинды и осенней семги в возрасте 3+1+ со структурой чешуи половозрелых пестряток. В результате мы пришли к выводу, что тинда — это карликовые особи, вернувшиеся в реку после года пребывания в море. А если их рассматривать по кратности нереста, то это их второй нерест после пребывания в море. Осенняя семга — это в прошлом неполовозрелые пестрятки и для них это будет первый нерест. Таким образом, различия в половой структуре рыб летнего и осеннего хода связаны с тем, что карликовые особи в реке представлены практически одними самцами.
Таблица 9 Размерная характеристика и темп роста карликовых самцов семги
|
Длина, |
Длина самцов по годам (L, см) и темп их роста |
|||||||
|
см |
0+ |
1+ |
2+ |
3+ |
||||
|
|
L |
T |
L |
T |
L |
T |
L |
T |
|
9,2 |
1,2 |
1,2 |
3,2 |
2,0 |
5,8 |
2,6 |
9,2 |
3,4 |
|
9,2 |
0,8 |
0,8 |
2,5 |
1,7 |
5,0 |
2,5 |
9,2 |
4,2 |
|
10,1 |
1,2 |
1,2 |
3,5 |
2,3 |
6,1 |
2,6 |
10,1 |
4,0 |
|
10,8 |
1,4 |
1,4 |
3,5 |
2,1 |
6,4 |
2,9 |
10,8 |
4,4 |
|
11,6 |
1,2 |
1,2 |
3,0 |
1,8 |
5,9 |
2,9 |
11,6 |
5,7 |
|
12,1 |
1,3 |
1,3 |
3,7 |
2,4 |
6,5 |
2,8 |
12,1 |
5,6 |
Наш вывод о разном происхождении тинды и осенней семги согласуется с мнением специалистов, которые пришли к такому же выводу на основании и непосредственных наблюдений (Нусенбаум, 1953; Митанс, 1973; Лейзерович, Мельникова, 1979; Schitffer, 1971).
Тинда в небольшом количестве представлена и самками — в р. Варзуга они, без учета возраста, в среднем составляют 1,9 % По аналогии с самцами, мы рассматриваем их как бывших половозрелых карликовых особей, идущих на второй нерест после пребывания в море. На это указывают схожесть структуры речной зоны чешуи; присутствие карликовых самок в естественных популяциях и появление их при искусственном разведении (Берг, 1961; Prouzet, 1981; Bagliniere, Massie, 1985; Hindar, Nordland, 1989; Яковенко, 1995); существование жилых популяций семги, состоящих только из карликовых самцов и самок, сформировавшихся в далеком прошлом (Dahl, 1927; Berg, 1953 и др.), и возврат в реку самок в год ската (Лысенко, 1997).
О биологических группах семги
На наличие в популяциях атлантического лосося рыб двух биологических групп обратил внимание в свое время Л. С. Берг (1943, 1948) который связал это явление с наличием у лосося двух рас — яровой и озимой. При этом тинду он относил к яровой расе, а рыб осеннего хода с неразвитыми половыми продуктами и с преобладанием самок — к озимой расе. Данное явление он связывал с разным действием температурного фактора на развитие половых продуктов. Однако многие исследователи имеют иную точку зрения об их происхождении.
Так В. В. Абакумов (1961) возникновение рас у лосося связывал с проходившим в антропогене вторжением и отступлением моря и меняющейся в этой связи протяженностью рек. При этом рыбы, нерестящиеся на дальних нерестилищах (озимая раса) должны были обладать большим запасом энергии, выносливостью, способностью выдерживать миграции на большие расстояния и продолжительное голодание. Рыбы, нерестящиеся на ближних нерестилищах (яровая раса), не совершали продолжительных миграций, поэтому имели меньше энергетических затрат.
По мнению Ю. П. Зелинского (1985) деление анадромных мигрантов лосося на две формы наследственно детерминировано, и существует такой механизм, который взрослым особям, находящимся в море, подает сигнал к прекращению нагула и началу анадромной миграции.
Гарднер (Gardner, 1976), обобщивший обширный материал по атлантическому лососю с начала столетия, отмечал, что о происхождении двух форм грилз (созревающие и не созревающие в момент захода в реку) существует две точки зрения. Одни исследователи связывают их появление с абиотическими факторами среды, другие усматривают в этом генетическую обусловленность. Но чем она вызвана, эти авторы, как и Ю. П. Зелинский, не объясняют.
Скарнекшина (Scarnecchina, 1983) связывал разное соотношение самцов и самок у рыб, проживших год в море, с протяженностью рек, широтой, местом нахождения реки, высотой склона, откуда стекает река, а также температурой воды в реке и в море в июле.
Таким образом, отметив наличие двух типов рыб у атлантического лосося, которые, при возвращении в реку после одного года пребывания в море, имеют разное состояние половых продуктов, исследователи объясняют по-разному. Кроме того, эти рыбы рассматриваются ими независимо друг от друга, т.е. как самостоятельные группы. Наши суждения о происхождении этих двух групп изложены в предыдущей главе. Исходя из этих предпосылок, рассмотрим более детально, как формируются тинда и осенняя семга с одним годом пребывания в море.
Карликовые особи, образующие впоследствии тинду, нерестятся первый раз, как правило, в возрасте 2+. После первого нереста и ската в море они возвращаются на следующий год в реку в летний период с развитыми половыми продуктами, которые в начале хода находятся во II-III, а в конце — IV-V стадиях зрелости. Таким образом, по кратности нереста для тинды — это второй нерест. Почему же они, в отличие от осенней семги, заходят в реку с развитыми половыми продуктами. Это становится понятным, если их сопоставить с теми осенними производителями, с которыми они отнерестовали в первый раз. У тех и других между первым и вторым нерестом проходит один год, однако первые проводят его в море, а вторые — в реке, У рыб, зимующих в реке после первого нереста, половые продукты находятся в состоянии вегетационного покоя, тогда как, находящиеся в это время в море карликовые самцы, активно питаются и растут, на следующий год у них происходит быстрое созревание половых продуктов и они вступают в новый нерестовый цикл. Таким образом, изначально разное поведение молоди в пресноводный период жизни наследственно обусловлено ходом эволюции атлантического лосося (Лысенко, 1994), что нашло свое отображение в поведении взрослых рыб в виде существования двух биологических групп. Способствующие быстрому созреванию карликовых самцов абиотические условия, являются лишь сопутствующим элементом. Это хорошо видно на примере тихоокеанских лососей. У большинства из них имеются карликовые особи, и исключение составляют лишь кета и горбуша. И какими бы абиотическими факторами на эти два вида не воздействовали, такая молодь у них в потомстве не образуется.
Половая структура нерестового стада
Установление родственных связей между двумя биологическими группами позволило затронуть еще один вопрос — каким было соотношение самцов и самок в родственной генерации в момент их рождения, независимо от того, какую в дальнейшем они формируют биологическую группу. Локальная популяция семги р. Варзуга является удобным объектом для такого исследования, поскольку, в обеих группах во время захода в реку доминирует одна возрастная группа — 3+1+, кроме того, к этой генерации относятся также рыбы в возрасте 2+1+ и 4+1+.
Если, например (табл. 10), 1974 г. — год возврата в реку основных осенних производителей, то 1975 г. — год их совместного нереста с тиндой другого поколения и карликовыми самцами, 1976 г. — год появления от них молоди в возрасте 0+, 1977 г. — возраст молоди 1+, 1978 г. — возраст молоди 2+ и ее скат в море, 1979 г. — возврат в реку рыб в возрасте 2+1+ и скат молоди в возрасте 3+, 1980 г. — возврат в реку рыб в возрасте 3+1+ в виде двух групп и скат молоди в возрасте 4+, 1981 г. — возврат в реку рыб в возрасте 4+1+. В пробе 1979 г. (потомство от осенних производителей 1974 г. возврата) в количестве 1939 экз. рыб в возрасте 2+1+ было 288 экз. Из них самцы и самки тинды составили 69,8 и 2,1%, а самцы и самки осенней семги -3,1 и 25,0 %. В 1980 г. в пробе из 1582 экз. рыб в возрасте 3+1+ насчитывалось 934 экз. самцов и самок тинды было, соответственно, 1,1 % и 10,7 %, а самок и самцов осенней семги — 61,0 % и 27,2 %. В 1981 г. в пробе из 1640 экз. только 4 рыбы имели возраст 4+1+, при этом они принадлежали к осенней семге, а количество самцов и самок было одинаковым. Суммарная проба рыб, вернувшихся в реку в возрасте 2+1+, 3+1+ и 4+1+ , равнялась 1226 экз. Самцы и самки тинды составили 24,6 и 1,3%, осенней семги соответственно — 21,6 и 52,5 %. По такому принципу был проанализирован весь материал за период 1979-1989 гг. В среднем в состав одной генерации входит 25,4 % самцов и 1,9 % самок тинды, 21,0 % самцов и 51,7 % самок осенней семги.
Таблица 10 Динамика полового состава нерестовых мигрантов
|
Год захода произво- |
Семга летнего хода в возрасте 2+1+, 3+1+, 4+1+ |
Семга осеннего хода в возрасте 2+1+, 3+1+, |
Численность поколения, |
||
|
дителей |
самцы, % |
самки, % |
самцы, % |
самки, % |
тыс. экз. |
|
1974 |
24,6 |
1,3 |
21,6 |
52,5 |
16,6 |
|
1975 |
19,1 |
1,5 |
27,7 |
51,7 |
15,4 |
|
1976 |
8,7 |
0,8 |
26,7 |
63,8 |
11,9 |
|
1977 |
19,1 |
1,3 |
23,2 |
56,4 |
23,4 |
|
1978 |
29,2 |
2,3 |
18,4 |
50,1 |
16,4 |
|
1979 |
28,2 |
2,5 |
20,4 |
48,9 |
20,2 |
|
1980 |
31,3 |
1,8 |
18,3 |
48,6 |
33,6 |
|
1981 |
30,5 |
3,0 |
18,0 |
48,2 |
50,4 |
|
1982 |
37,7 |
2,6 |
14,6 |
45,2 |
38,6 |
Представленные данные показывают, что при увеличении численности стада возрастает доля самцов тинды и снижается доля осенних самцов. Так, от числа производителей 1974 г. возврат самцов тинды составил 24,6 %, а самцов осенней семги — 21,6 %, от числа же производителей 1980 г., соответственно, 31,3 % и 18,3 %. Кроме того, при увеличении численности стада снижается количество самок осенней семги, а при уменьшении — доля их возрастает. Если не учитывать принадлежность особей к одной из групп, то соотношение самцов и самок близко к 1:1.
Семга реки Кица — притока реки Варзуга
Река Кица впадает в реку Варзуга всего в 4-х км от моря в районе обширной эстуарной зоны (рис. 1 ) и, как нам представляется, именно это обстоятельство оказало основное влияние на степень проявления инстинкта хоминга у семги, что, со временем, привело к формированию отдельных ее субпопуляций в этих реках. О реальности подобных утверждений свидетельствуют, в частности, скат анадромных варзугских мигрантов, зашедших «по ошибке» в р. Кица на приливе, что подтверждено мечением (В. А. Бухтияров — неопубликованные данные 1970-1971 гг.), и некоторые особенности строения чешуи, которые позволяют достаточно объективно разделять рыб на варзугских и кицких.
Динамика хода нерестовых мигрантов
Ход семги в реку начинается после окончания ледохода и заканчивается либо в конце ноября, либо в течение декабря (табл. 11).
Таблица 11 Сроки хода разных биологических групп семги в р. Кица
|
биологические группы |
сроки хода |
| заледка | середина-конец мая — начало июня |
| закройка | середина июня — конец июля |
| тинда | конец июня — начало августа |
| осенние мигранты | середина августа — декабрь |
Первыми в реку заходят заледка и закройка, затем тинда, и завершают ход осенние мигранты. Однако в зависимости от гидрологических условий реки сроки миграций могут сдвигаться на одну неделю и более. Такое, например, было в 1979, 1994 и 1997 гг., когда тинду стали ловить в начале июля, а осеннюю семгу — в начале сентября. Заледки и закройки в ходовом стаде мало, иногда закройка может отсутствовать вовсе.
Характеристика семги р. Кица
Основу ходового стада семги р. Кица, как и р. Варзуга, составляют рыбы в возрасте 3+1.
Тинда представлена тремя возрастными группами — 2+1+, 3+1+, 4+1+ — и на 98 % состоит из самцов, преимущественно в возрасте 3+1+. В среднем тинда, без учета возраста и пола, достигает длины 53,6 см и массы 1,66 кг (табл. 12). Минимальная длина — 39 см — была у самца в возрасте 2+1+. Тинда в возрасте 5+1+ встречается в р. Кица крайне редко, при этом самки имеют длину 55-59 см, а самцы, соответственно, 58-63 см.
Плодовитости самок кицкой тинды в возрасте 2+1+ и 3+1+ составляет 2,6-5,2 тыс. икринок (табл. 13), в среднем 3,8 тыс. икринок.
Таблица 13 Плодовитость семги р. Кица
|
|
|
Длина, |
Плодовитость, тыс. шт. икринок |
|||||
|
Возраст |
N, |
см |
индивидуальная |
относительная |
||||
|
|
экз. |
пределы |
средняя |
пределы |
средняя |
пределы |
средняя |
|
|
тинда |
||||||||
|
2+1+ |
12 |
45-53 |
48,1 |
2,6-4,0 |
3,4 |
1,9-3,1 |
3,0 |
|
|
3+1+ |
30 |
45-55 |
50,0 |
3,3-5,2 |
4,0 |
2,5-3,4 |
3,0 |
|
|
осенняя семга |
||||||||
|
3+1+ |
40 |
55-65 |
57,0 |
6,1-9,6 |
6,8 |
2,7-5,4 |
3,5 |
|
|
3+2+ |
32 |
63-74 |
66,0 |
7,1-12,2 |
8,9 |
2,9-4,1 |
3,2 |
|
Осенние мигранты представлены в основном пятью возрастными группами — 2+1+, 3+1+, 4+1+, 2+2+ и 3+2+ и на 70 % состоит из самок, преимущественно в возрасте 3+1+, в небольшом количестве встречаются рыбы в возрасте 4+2+ и совсем редко — 5+1+. При этом минимальная длина самцов в возрасте 3+1+ составляла 52 см, а максимальная 75 см, а самок того же возраста, соответственно 48 и 72 см. Длина рыб в возрасте 5+1+ колебалась от 58 до 62 см. За период наших наблюдений наибольшую длину имел самец в возрасте 3+2+ — 90 см.
Количество повторно нерестующих рыб у кицкой семги меньше, чем у варзугской, и составляет в среднем 1,6 %. Длина этих рыб 68-84 см, масса — от 3,6 до 6,0 кг. Примечательно, что в р. Кица была поймана семга в возрасте 5+1+Sm длиной 65 см. Особи с таким возрастом в р. Варзуга нам не попадались. Особенностью кицкой семги можно также считать и то, что она может возвращаться на повторный нерест не только в год ската, но и летом следующего года, чего не наблюдается у варзугских рыб.
Плодовитости рыб осеннего хода в возрасте 3+1+ и 3+2+ составляет 6,1-12,2 тыс. икринок (табл. 13), в среднем 8,0 тыс. икринок.
Средняя длина кицкой семги составляет 57,6 см, а масса 2,5 кг. Соотношение самцов и самок в ходовом стаде в среднем составляет 41,0 и 59,0 %, а в нерестовом стаде оно несколько иное — 38,0 % и 62,0 %.
Возрастная структура нерестового стада
Возрастная структура нерестового стада р. Кица (табл. 14) была исследована в период 1975-1990 гг. Как видно из приведенных данных, она практически не отличается от таковой по субпопуляции семги р. Варзуга.
Численность семги р. Кица и причины ее колебаний
Численность ходового стада семги в р. Кица в 1975-1990 гг. колебалась от 2,2 до 13,7 тыс. экз. (табл. 15), т.е. она на порядок ниже, чем в р. Варзуга. С учетом того, что при определенных климатических и гидрологических условиях в эстуарную зону р. Кица через РУЗ заходит во время прилива варзугская семга, которая затем скатывается, доля непосредственно кицких мигрантов еще меньше
Таблица 15 Характеристика стада семги р. Кица и уровень его эксплуатации за период 1975-90 гг.
|
Год |
Числен-ность, |
Вылов, |
Ср. масса особи, |
Соотношение полов, % |
Соотношение рыб, % |
|||
|
|
тыс. экз. |
тыс. экз. |
ц |
кг |
самцы |
самки |
тинда |
осенняя |
|
1975 |
13,7 |
6,7 |
159 |
2,3 |
55,3 |
44,7 |
23,5 |
76,5 |
|
1976 |
7,8 |
4,3 |
115 |
2,7 |
41,8 |
58,2 |
29,1 |
70,9 |
|
1977 |
8,2 |
4,2 |
93 |
2,2 |
52,5 |
47,5 |
14,4 |
85,6 |
|
1978 |
4,3 |
2,0 |
51 |
2,5 |
53,7 |
47,3 |
22,0 |
78,0 |
|
1979 |
5,8 |
2,8 |
70 |
2,5 |
42,9 |
57,1 |
5,3 |
94,7 |
|
1980 |
2,2 |
1,1 |
27 |
2,5 |
51,4 |
48,6 |
45,8 |
54,2 |
|
1981 |
4,3 |
2,1 |
54 |
2,5 |
41,5 |
58,5 |
6,7 |
93,3 |
|
1982 |
4,0 |
2,0 |
54 |
2,8 |
37,2 |
62,8 |
13,6 |
86,4 |
|
1983 |
3,6 |
1,7 |
41 |
2,4 |
44,4 |
55,6 |
33,6 |
66,4 |
|
1984 |
7,1 |
3,4 |
96 |
2,9 |
35,5 |
64,5 |
11,6 |
88,4 |
|
1985 |
10,7 |
5,1 |
149 |
2,9 |
40,5 |
59,5 |
16,1 |
83,9 |
|
1986 |
6,9 |
3,3 |
86 |
2,6 |
39,2 |
60,8 |
20,7 |
79,3 |
|
1987 |
9,7 |
3,5 |
93 |
2,7 |
35,4 |
64,6 |
12,4 |
87,6 |
|
1988 |
10,4 |
3,8 |
92 |
2,4 |
43,6 |
56,4 |
19,0 |
81,0 |
|
1989 |
3,5 |
1,3 |
32 |
2,5 |
58,9 |
41,1 |
50,8 |
49,1* |
|
1990 |
6,2 |
2,3 |
58 |
2,5 |
44,4 |
55,6 |
35,1 |
64,9* |
Минимальное количество тинды в этот период было 0,3 тыс. экз., максимальное — 2,1 тыс., в среднем — 1,2 тыс., а семги осеннего хода, соответственно, 1,3 тыс., 10,0 тыс. и 5,1 тыс. Соотношение количества тинды и семги осеннего хода в среднем составляло 1:4,1.
В р. Кица, как и в р. Варзуга, существует шесть основополагающих стад (линий) семги, каждому из которых свойственен шестилетний цикл оборачиваемости поколений. На основании анализа материалов за 1966-1995 гг. цикл оборачиваемости поколений, если синхронизировать его с данными по варзугской популяции семги, можно представить так:
1966-1972-1978-1984-1990 — 1 линия
1967-1973-1979-1985-1991 — 2 линия
1968-1974-1980-1986-1992 — 3 линия
1969-1975-1981-1987-1993 — 4 линия
1970-1976-1982-1988-1994 — 5 линия
1971-1977-1983-1989-1995 — 6 линия
Графическое изображение динамики хода с выделением в нем шести воспроизводящихся линий (рис. 4) позволяет наглядно показать закономерности чередования спадов и подъемов численности, как по отдельным линиям, так и в стаде в целом.
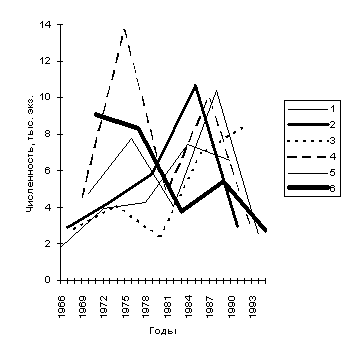
Рис. 4. Динамика численности ходового стада семги р. Кица за период 1966-1995 гг.
Если сравнивать динамику хода семги в реки Варзуга и Кица, то спады и подъемы численности стад приходятся в основном на одни и те же годы. Таким образом, схожесть многих черт биологии семги этих рек при их независимом существовании свидетельствует о том, что формирование этих популяций шло по единому природному сценарию.
Ход горбуши в реки Варзуга и Кица
В 1956 г. было подготовлено биологическое обоснование акклиматизации горбуши в бассейне Белого и Баренцева морей, которое послужило основанием для принятия решения о начале этих работ, которые, с перерывами, продолжаются до сих пор.
До 1997 г. наибольший возврат горбуши, по учетным данным, наблюдался в 1973 г. — 262 тыс. экз. Начиная с 1985 г. горбуша в массе заходила в реки Кольского п-ова только в нечетные годы, хотя раньше наблюдался ее возврат и в четные годы — например, в 1974 г. При этом численность ее последнее время постоянно увеличивается. Как правило, большая часть производителей горбуши мигрировала в малые реки Кольского п-ова. В таких крупных реках, как Варзуга, фиксировался проход на нерест от нескольких десятков до нескольких тысяч рыб. Однако в 1997 г. наблюдался наибольший по численности заход горбуши в реки Варзуга и Кица (табл. 16)
Таблица 16 Статистика хода горбуши в реки Варзуга и Кица и уровень ее вылова
|
|
Варзуга |
Кица |
|||||||||
|
Год |
учтено, |
пропуск, |
сдано |
учтено |
пропуск, |
сдано |
|||||
|
|
экз. |
экз. |
экз. |
ц |
экз. |
экз. |
экз. |
ц |
|||
|
1987 |
Всего учтено11 экз. |
||||||||||
|
1989 |
101 |
- |
101 |
- |
143 |
90 |
42 |
0,8 |
|||
|
1991 |
591 |
372 |
165 |
2,7 |
1119 |
765 |
394 |
6,3 |
|||
|
1993 |
3220 |
2028 |
1101 |
18,8 |
969 |
609 |
334 |
5,5 |
|||
|
1995 |
3785 |
2153 |
1339 |
20,4 |
6804 |
4286 |
2480 |
36,9 |
|||
|
1997 |
18392 |
1072 |
17183 |
243,3 |
8781 |
2000 |
6441 |
80,1 |
|||
В связи с этим представляется важным рассмотреть возможные причины, вызвавшие вспышку численности горбуши и обсудить, какое влияние оказал массовый ход горбуши на динамику и миграционное поведение тинды.
В р. Варзуга в 1997 г. первые особи горбуши были отмечены в уловах 2 июля, и ход ее продолжался до 5 августа. Пик наблюдался 20 июля — в этот день было учтено 5,8 тыс. рыб общим весом 87 ц. Первые особи горбуши в р. Кица были отмечены 2 июля, однако ход ее здесь был более продолжительным — до 12 августа. Вылов горбуши в р. Варзуга составил 17,2 тыс. экз. или 245 ц, общая ее численность оценена в 18,4 тыс. экз. или 262 ц, а в р. Кица, соответственно, 6,4 тыс. экз. или 81 ц и 8,8 тыс. экз. и 110 ц. Длина рыб в уловах колебалась от 30,0 до 50,6 см, а масса — от 0,37 до 3,0 кг.
Достаточно большими были уловы горбуши и на морских тонях, расположенных вдоль всего Терского берега. Так, на морской тоне у села Чаваньга с 8 по 25 июля было добыто 20,8 тыс. экз. — 285 ц, а у села Чапома с 23 июня по 25 июля, соответственно, 26,2 тыс. экз. — 359 ц.
Большой подъем численности горбуши можно объяснить лишь тем, что в 1995 г. в период ее нереста и развития икры до устойчивой стадии — с третьей декады августа до первой декады октября — температурный режим был крайне благоприятным и средняя декадная температура воды в р. Варзуга составляла, соответственно, 10,8, 11,2, 7.0, 7,7 и 5.9 0С. Кроме того, следует учесть, что в 1996 г. Умбским и Кандалакшским рыбоводными заводами было выпущено, соответственно, 502 тыс. и 51 тыс. личинок горбуши. По-видимому, в совокупности эти два фактора и явились причиной возврата мощного по численности поколения горбуши.
А как такой массовый подход горбуши отразился на миграции тинды? Проводившиеся исследования показали, что при малой численности горбуши в предыдущие годы, ход летней рыбы начинался раньше — в последней декаде июня. В 1997 г. тинда в обеих реках была отмечена только в первой декаде июля и на протяжении первой половины июля численность летней рыбы была незначительной. Только в конце июля, когда миграция горбуши стала заканчиваться, начался активный ход летнего лосося, который продолжался до конца августа, обычно же он заканчивается в первой его декаде. Отметим, что в этих реках наблюдается задержка хода летней рыбы, связанная с температурным фактором, однако, в данном случае, если он и сыграл свою роль в сроках начала миграции летней рыбы (температура воды в р. Варзуга в конце июня-начале июля была около 180С, что примерно на 4 0С выше, чем среднемноголетняя), то существенная задержка массового хода тинды была отмечена нами впервые и, вполне возможно, обусловлена именно массовым ходом горбуши.
Летняя семга, мигрировавшая в реку в 1997 г., сильно отличалась от обычной тинды. Обычно тинда в начале хода хорошо упитана, пиларические придатки покрыты слоем жира, половые продукты находятся во II-III стадиях зрелости, печень в нормальном состоянии. А эта рыба по виду была больше похожа на подлошавшую тинду в конце хода — прогонистая и тощая, на пилорических придатках почти нет жира, печень тощая, но вот половые продукты еще лишь во II-III стадиях зрелости. Среди них попадались особи, у которых структура речной зоны чешуи была отличной от таковой варзугской семги. Данный факт наводит на мысль о заходе в реку “чужаков”, которые возможно были занесены в реку мощным потоком горбуши. Только в конце июля, когда почти закончился ход горбуши, стала появляться тинда с обычными показателями своего состояния.
А как массовый ход горбуши на нерест может повлиять на стоящую на нерестилищах семгу? Таких сведений в литературе нет, и мы ими также пока не располагаем. Но в 1995 г., например, когда в р. Кица было пропущено 4,3 тыс. экз. горбуши, на нерестилищах в это время находилось 1,2 тыс. экз. осенней семги с предыдущего года. О результатах их взаимоотношений можно будет судить, видимо, только в 2000 г.
В связи с рассматриваемыми вопросами, следует представить сведения о необычном лососе (рис.5), пойманном в ноябре 1995 г. Этот экземпляр был передан нам в выпотрошенном виде. Как видно на снимке, задняя часть тела у него типичная семужья, а вот передняя — как у лошающей горбуши. Гибрид это или урод — установить не удалось, но нам такая рыба попалась впервые. По возрасту — 3+1+ — ее, разумеется, следует отнести к семге. Хотя других сведений у нас нет, мы полагаем, что этот факт заслуживает внимания. Если это урод, то данное обстоятельство может служить индикатором ухудшающийся экологической обстановки, что отмечено в 1991 г., когда среди мигрирующей семги было много рыб с какими-то ржавыми пятнами по всему телу. А может быть это — результат биологического загрязнения рек? В связи с чем, уместно заметить, что есть достоверные сведения о случаях захода в малые реки Восточного Мурмана стальноголового лосося Salmo gaidneri Richardson, сбежавшего, очевидно, с рыбоводных ферм соседних стран.

Рис. 5. Лосось, пойманный в устье р. Варзуга в ноябре 1995 г.
Появление большого количества горбуши в самой крупной лососевой реке, какой является Варзуга, настораживает и по другой причине. Как известно современная техногенная деятельность человека привела к тому, что условия обитания лососевых рыб ухудшаются, в результате чего происходит уменьшение речных площадей, пригодных для их обитания. В связи с этим горбуша может вытеснить более малочисленного конкурента. Поэтому мы солидарны с Ю. П. Алтуховым (1997), который считает, что так называемая успешная акклиматизация лососей, часто приводит к угнетению ценнейших аборигенных популяций. На подобные факты обращает внимание и Международный Союз по сохранению природы и природных ресурсов, который рассматривает акклиматизацию организмов в новых местах как форму биологического загрязнения.
Подводя итог вышесказанному, считаем — для того, чтобы обезопасить крупнейшую лососевую реку Кольского полуострова от такого массового биологического загрязнителя, каким может стать акклиматизация горбуши, необходимо полностью изымать ее на РУЗ.
Заключение
Многолетние непрерывные наблюдения за популяцией атлантического лосося р. Варзуга позволили, помимо традиционных сведений о длине, массе, половом и возрастном составе, сроках хода анадромных мигрантов, получить новые данные о биологии семги этого стада. Проведение аналогичных исследований семги р. Кица, которые в таком объеме публикуются впервые, позволило не только составить представление о ее биологии, но и выявить много общего между стадами лосося двух рек, которые, предположительно, практически не смешиваются друг с другом.
Сроки хода в реку варзугской и кицкой семги совпадают. Началом их миграции служит окончание ледохода. Первыми заходят заледка и закройка, затем начинается ход тинды, которая возвращается в р. Кица несколько раньше, чем в р. Варзуга, и ход ее оканчивается на неделю раньше хода варзугской тинды.
У семги р. Варзуга между летним и осенним ходом почти нет перерыва, тогда как в р. Кица такой перерыв в ходе может составлять от двух недель до месяца.
Основу двух биологических групп формируют рыбы в возрасте 3+1+, что свидетельствует об их принадлежности к одной генерации. Летняя форма (тинда) у лососей обоих рек на 98,0 % представлена самцами, а осенняя — на 70,0 % самками. Им свойственен почти одинаковый процент повторно нерестующих особей в ходовом стаде — так у варзугской семги он составляет в среднем 1,9 %, а у кицкой семги — 1,6 %. В популяциях семги обоих рек наблюдается одинаковая зависимость — увеличение численности стада происходит всегда за счет рыб с абсолютным возрастом 4+, и, при этом, рыба оказывается в целом мельче.
Положенный в основу анализа динамики хода позволил установить, что для популяций семги обеих рек характерно наличие шести основных репродуктивных стад, а колебания численности в каждой воспроизводящей линии, несмотря на то, что для исследования использованы данные по динамике хода за разное количество лет, подчинены одной и той же закономерности — чередование спадов и подъемов с интервалом в 6 лет. При этом спады и подъемы численности обоих стад приходятся в основном на одни и те же годы. Проведенный сравнительный анализ биологии семги рек Варзуга и Кица показывает, что эволюционный процесс проходил здесь по единому природному сценарию.
Использование чешуи половозрелых и неполовозрелых пестряток в качестве эталона для идентификации анадромных мигрантов позволило установить, что тинда обоих полов — это повторно нерестующие, после пребывания в море, бывшие половозрелые пестрятки (карликовые особи). Рыбы осеннего хода — это первый раз нерестящиеся, после пребывания в море, бывшие неполовозрелые пестрятки.
Сопоставление цикла развития тинды от молоди до взрослой рыбы с циклом развития взрослых рыб от молоди до возвращения их на повторный нерест дало возможность объяснить, почему самцы тинды оплодотворяют самок не своего, а другого поколения.
Подробный анализ возрастной структуры нерестового стада варзугской семги позволил выяснить качественный состав особей, принимающих участие в нересте. Установлено, что в нересте принимают участие 8 поколений, составляющих 12 возрастных групп, из которых основными являются рыбы в возрасте 3+1+, 3 возрастные группы тинды и 3 возрастные группы карликовых самцов. Длительные наблюдения позволили обнаружить самок, вернувшихся в реку после нереста в год ската.
Статистика хода и вылова горбуши, особенно за 1997 г., свидетельствует о том, что численность этого акклиматизированного лосося в настоящее время возрастает, и он начинает в массе заходить в реки Варзуга и Кица. Это не может не вызывать беспокойство за сохранность ценнейших популяций семги, поэтому следует обезопасить эти реки от возможных негативных последствий подобной экспансии, для чего необходимо максимальное изъятие горбуши на РУЗ.
Литература
Абакумов В. В. Сезонные расы проходных рыб // Вопр. ихтиологии, 1961. Вып. 17. С.179-190.
Азбелев В. В. Материалы по биологии семги Кольского полуострова и ее выживаемость // Тр. ПИНРО, 1960. Вып. 12, С. 5-70.
Азбелев В. В. Семга Баренцева моря // Рыбы Мурманской области. Мурманск, 1966. С.169-176.
Азбелев В. В. Численность и использование семги рек Кольского полуострова // Тр. ПИНРО. 1968. Вып.23. С.513-526.
Азерникова О. А. Метод расчета численности семги реки Печоры // Рыб. хоз-во. 1964. №2. С.24-43.
Алтухов Ю. П., Варнавская Н. В. Адаптивная генетическая структура и ее связь с внутрипопуляционной дифференциацией по полу, возрасту и скорости роста у тихоокеанского лосося — нерки, Oncorhynchus nerka (Walbaum) // Генетика. 1983. Т.19. №3. С.796-807.
Алтухов Ю. П., Салменкова Е. А. Омельченко В. Т. Популяционная генетика лососевых рыб. М.: Наука, 1997.
С.223-225.
Бакштанский Э. Л., Яковенко М. Я. Миграции вальчаков атлантического лосося из р. Варзуга // Тр.
ВНИРО. 1976. Т.116. С.33-38.
Берг Л. С. Яровая и озимая раса у проходных рыб // Изв. АН СССР / Отд. физ.-мат. и естеств. Наук. 1934. №5.
С.711-732.
Берг Л. С. Избранные труды. М.-Л., 1961. С.475-666.
Бирман И. Б. Лососевые и солнечная активность // Тр. ПИНРО. 1969. Т.67. С.171-189.
Бухтияров В. А. Лысенко Л. Ф. Характеристика стада семги реки Варзуги и рекомендации по рационализации
промысла // Рукопись / ПИНРО. 1970. 32 с.
Бухтияров В. А., Лысенко Л. Ф. Характеристика стада семги реки Варзуги хода 1971 г. и перспективы увеличения ее численности // Рукопись / ПИНРО. 1971. 60 с.
Горшкова Г. В., Горшков С. А. Карликовая нерка Oncorhynchus nerka в Азабачьем озере на Камчатке // Вопр. ихтиологии. 1977. Т.17. Вып.2. С.220-224.
Дерюгин К. М. Фауна Белого моря и условия ее существования // Исследование морей СССР. 1928. Вып.7-8.
С.338.
Драганов М. А., Мартынов В. Г., Лысенко Л. Ф. Условия естественного воспроизводства и популяционная структура атлантического лосося в бассейне реки Варзуги // Тр. Коми НЦ УРО АН СССР. 1990. №114. С.5-30.
Зелинский Ю. П. Структура и дифференциация популяций и форм атлантического лосося. Л.: Наука, 1985.
С.12-13.
Казаков В. Р. Продуцирование спермы карликовыми самцами атлантического лосося и характеристика ее
рыбоводных качеств. // Тр. ГосНИОРХ. 1979. №39. С.40-47.
Казаков В. Р., Кузьмина О. Т., Шустов Ю. А., Щуров И. Л. Атлантический лосось реки Варзуги. С-Пб:
Гидрометеоиздат, 1991. 107 с.
Лейзерович Х. А. О карликовых самцах при заводском разведении атлантического лосося // Вопр.
ихтиологии. 1973. Т.13. Вып.3. С.460-470.
Лейзерович Х. А., Мельникова М. Н. Миграции в море карликовых самцов атлантического лосося, Salmo
salar // Вопр. ихтиологии. 1979. Т.19. Вып.1. С.178-181.
Лившиц М. Г. Причины массовой гибели семги в р. Варзуга в 1969 г. // Науч.- тех. бюлл. ПИНРО. 1962.
№4(22).
Лысенко Л. Ф. О причинах колебания численности семги реки Варзуги // Вопросы лососевого хозяйства на
Европейском Севере. Петрозаводск, 1987. С.14-20.
Лысенко Л. Ф. О темпе роста тинды и осенних производителей семги реки Варзуги // Проблемы изуч., рац.
использ. и охраны природ. ресурсов Белого моря. / Тез. докл. регион. конф. Архангельск, 1990. С.168-170.
Лысенко Л. Ф. О тинде реки Варзуги и причинах колебания ее численности // Проблемы изуч., рац. использ.
и охраны природ. ресурсов Белого моря. / Тез. докл. регион. конф. Архангельск, 1990. С.170-172.
Лысенко Л. Ф. Способ идентификации анадромных мигрантов атлантического лосося // а.с. 31655416, СССР.
МКМ5А01К61/100.-14684758/13. Бюл. №22. опубл.12.06.91.
Лысенко Л. Ф. Почему самцы тинды семги оплодотворяют самок осеннего хода другого поколения//
Проблемы изуч., рац. использ. и охраны природ. ресурсов Белого моря. / Тез. докл. регион. конф. Петрозаводск,
1992. С.182-183.
Лысенко Л. Ф. Массовая гибель семги в р. Варзуга // Рыбное хоз-во. 1994. №4. С.32.
Лысенко Л. Ф. Особенности половой структуры стада лосося на примере р. Варзуга // Материалы отчетной сессии по итогам НИР ПИНРО в 1993 г. / Изв. ПИНРО. 1994. С.245-256.
Лысенко Л. Ф. Атлантический лосось: биология и происхождение // Препринт. / Мурманск, 1994. 44 с.
Лысенко Л. Ф. Биологические особенности самок тинды атлантического лосося // Проблемы изуч., рац. использ. и охраны природ. ресурсов Белого моря. / Тез. докл. регион. конф. Санкт-Петербург, 1995. С.533-534
Лысенко Л. Ф. О структуре популяции атлантического лосося Salmo salar L. реки Варзуги (Кольский
полуостров) // Вопр. ихтиологии. 1997. Т.47. Вып.6. С.475-482.
Лысенко Л. Ф. О самках атлантического лосося Salmo salar L. , возвращающихся в реку в год ската
// Вопр. ихтиологии. 1977. Т.37. Вып.6. С.427-428.
Мартынов В. Г. Семга уральскихпритоков Печоры. Л.: Наука, 1983. 127 с.
Мартынов В. Г., Лысенко Л. Ф. Структура уловов в Варзугском промысловом районе // Проблемы изуч. и рац.
использ. природ. ресурсов Белого моря. / Тез. докл. регион. конф. Архангельск, 1985. С.249-250.
Мельникова М. Н. Опыт определения численности семги р. Варзуга // Науч. тех. бюлл. ВНИОРХ. 1958. №6-7.
С.27-32.
Мельникова М. Н. Биология семги р. Варзуга // Изв. ВНИРО. 1959. Т.48. С.132-150.
Мельникова М. Н. Об определении плодовитости семги реки Варзуги // Науч.- тех. бюлл. ВНИОРХ. 1962. С.27-28.
Мельникова М. Н. Плодовитость семги (Salmo salar) реки Варзуги // Вопр. ихтиологии. 1964. Т.3. Вып.3. С.469-476.
Мельникова М. Н. Семга Терского берега // Рыбы Мурманской области. Мурманск, 1966. С.152-168.
Мельникова М. Н., Персов П. М. О мечении молоди семги р. Варзуга и лосося на р. Нева // Тр. Кар.
ГосНИИОРХ. 1968. Т.5. Вып.2.С.81-83.
Мельникова М. Н. Некоторые особенности молоди семги (Salmo salar) ряда рек бассейна Белого моря //
Вопр. ихтиологии. 1970. Т.10. Вып.3. С.42-45.
Митанс А. Р. О карликовых самцах и половой структуре популяции балтийского лосося // Вопр. ихтиологии. 1973. Т.13. Вып.2. С.231-237.
Монастырский Г. Н. О состоянии сырьевых ресурсов семги Северного промышленного района // Тр. ВНИРО. 1935. Т.2. С.5-59.
Нусенбаум А. М. Исследование покатной молоди семги // Рыбное хоз-во. 1953. №9. С.21-25.
Плохинский Н. А. Биометрия. Новосибирск, 1961. 346 с.
Песлак Я. К. Влияние качества производителей на качество выращенной на рыбоводных заводах молоди семги //
Обмен веществ и биохимия рыб. М.: Наука, 1967. С.73-75.
Падецкий С. Н., Флоряк В. С. Особенности биологии симы Приморья в речной период жизни // Морфология,
структура популяции и проблемы рац. использ. лососевид. рыб. Л., 1983. С.89-90.
Покровский В. Р. О причинах колебания численности семги Salmo salar // Тр. Кар. ГосНИИОРХ. 1966. Т.4.
Вып.1. С.19-26.
Салмов В. З. Оценка условий естественного воспроизводства семги // Биологические ресурсы Белого моря
и водоемов Европейского Севера. Петрозаводск, 1981. С.102-107.
Якобсон Р. П. Отчет по обследованию рыболовных угодий Александровского и Кемского уездов Архангельской
губернии // Материалы к познанию русского рыболовства. 1914. Т.3. Вып.2. 115 с.
Bagliniere J.J., Massie G.Precocious and maturation in wild Atlantic salmon in the Armorica France // Aquaculture. 1985. V. 45. №1-4. P.259-253.
Bailey J.K., Saundres R. J., Buzeta M.J. Influence of parental smolt and sea age to growth and smolting of hatchery-reared Atlantic salmon (Salmo salar) // Can. J. Fish. Aquat. Sci. 1980. V.37. №6. P.1379-1386
Berg M.A. A relict Salmo salar called “smablank” from river Namsen // North Trondelag. Acta Borealia. Tromso. Musuem, 1953. №6.
Dahl K. The «blege» or «Dwarf» salmon // A landlocked from lake Byglandsfjord in setesdal / Skr. Nor. Vid. ensk. Akad. Oslo, 1927. №9. 25 p.
GardnerM.L.G. A review of factors which influence the sea-age and maturation of Atlantic salmon, Salmo salar L. // J. Fish. Biol. 1976. V.9. №4. P.229-237.
Hindar K., Nordland J. A. A female Atlantic salmon (Salmo salar) maturing in the parr stage. // J. Fish. Biol. 1989. V.35. №3. P.461-463.
Jones J.W. Studies on the scales of young salmon (Salmo salar) jiv. in relation to growth, migration and spawning // Fish. investigation. 1949. Ser.1. V.1. 23 p.
Larsson P. O., Svensson K. M. Studies on the possible influence of early maturation grilse frequency by means of tapping experiments in the river Zuie // ICES. C.M. m., 1974. 28. 7 p.
Myres R. A. Hutchingst J. A., Gibson R. J. Variation on male parr maturation and population of Atlantic salmon (Salmo salar) // Can. J. Fish. Aquat. Sci. 1986. V.43. №6. P.1242-1248.
Prouzet P. Observation d’una femelle de tacon de saumon atlantiqu (Salmo salar) parvenue a maturite sexuelle an riviere // Bull. Franc. piscicult. 1981. V.54. №282. P.16-19.
Major R.J., Masher K.H., Mason J.E. Identification of stock of Pacific salmon by means of scale features // MacMilan Lectures / Univ. British Columbia. 1972. P.209-228.
Scarnecchina D.L. Age at sexual maturity in Icelandic stocks of Atlantic salmon (Salmo salar) // Can. J. Fish. Aquat. Sci. 1983. V.40. №9. P.1450-1468.
Schieffer K. Ecology of Atlantic salmon with special reference to occurrence and abundance of grilse in North Shire of Lawrence River // PhD. Thesis Univ. of Waterloo. 1971. 129 p.
Shaffer M., Elson P. J. Adaptive significance of variation in life history among local of Atlantic salmon in North American // Ecology. 1975. №56. P.571-590.
Thorpe J. E., Morgan R. J., Talbot C., Miles M. S. Inheritance of development rates in Atlantic (Salmo salar) // Aquaculture. 1983. V.33. №1-4. P.119-128.
Thomaz D., Beall E., Burke T. Alternative reproduction tactics in Atlantic salmon: Factors affecting mature parr success // Proc. Roy. Soc. London. B. 1997. V.264. №1379. P.219-226.